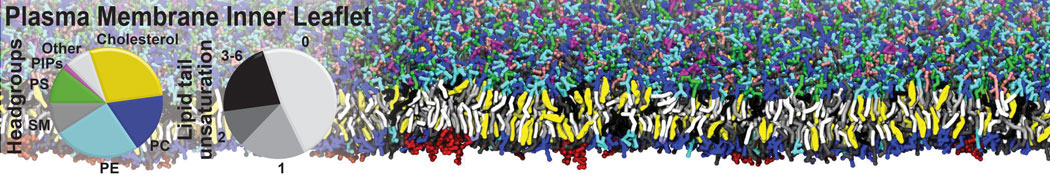
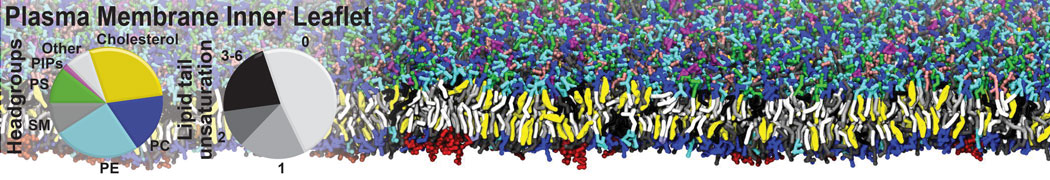
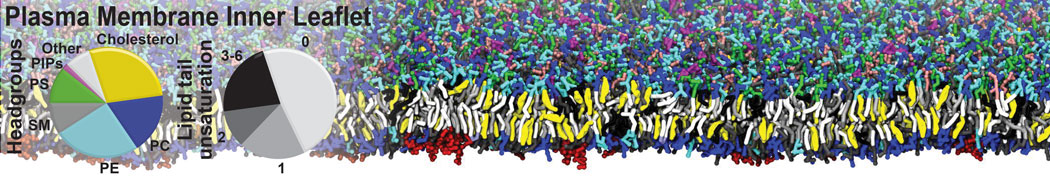
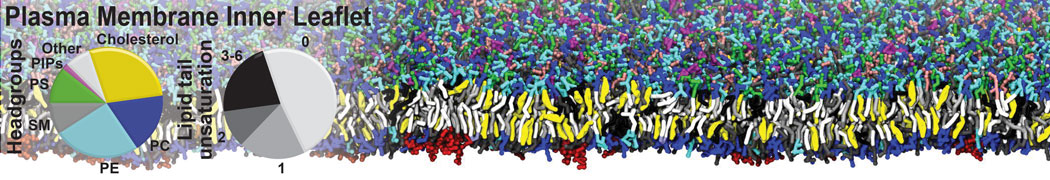
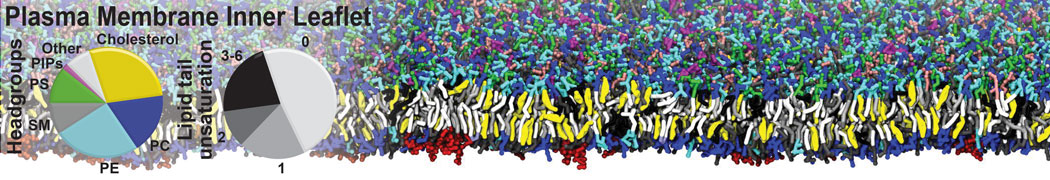
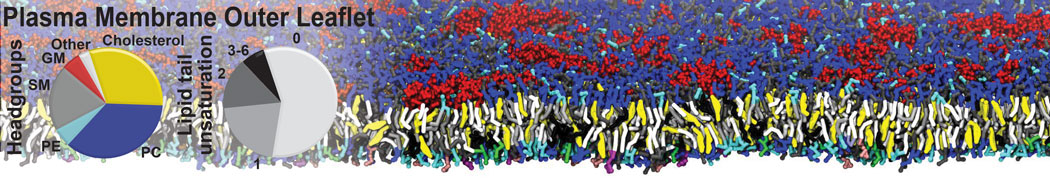
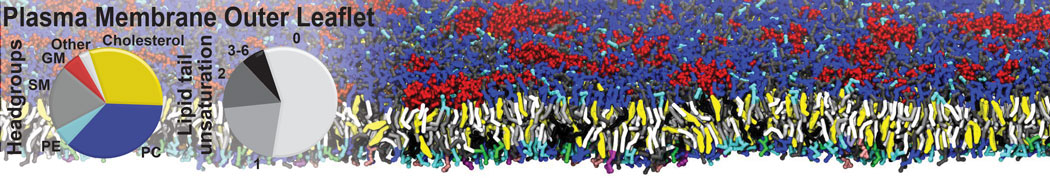
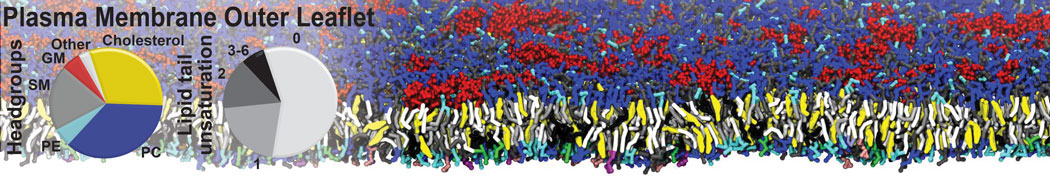
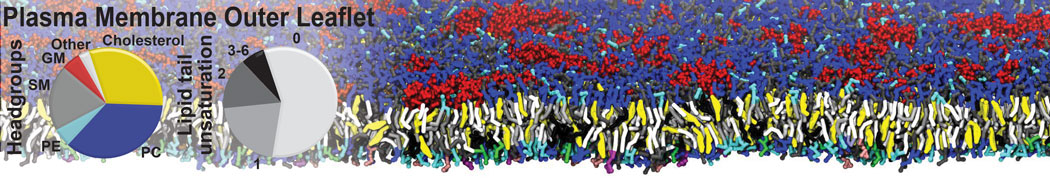
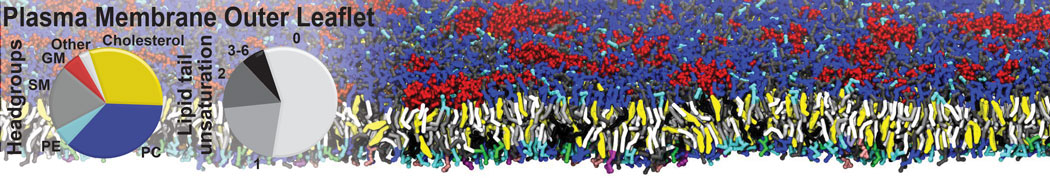
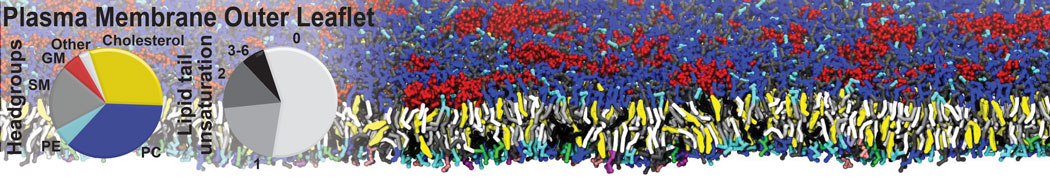
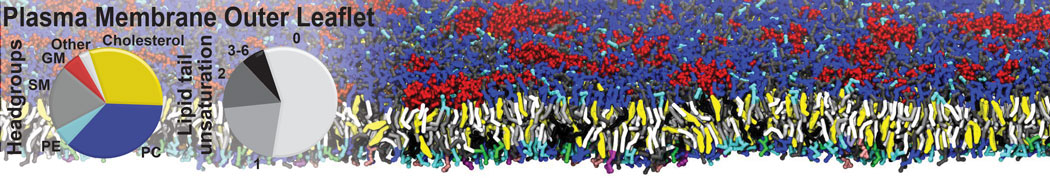
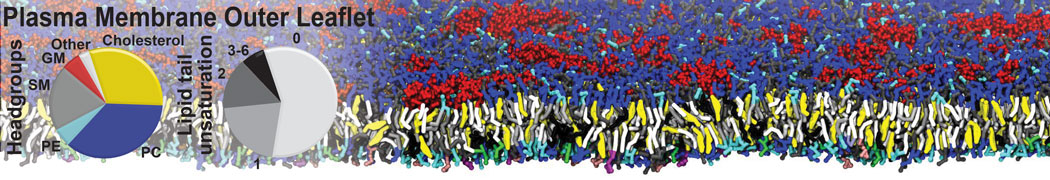
Raft & Amphiphiles
- Details
-
Last Updated: Wednesday, 21 August 2013 11:44
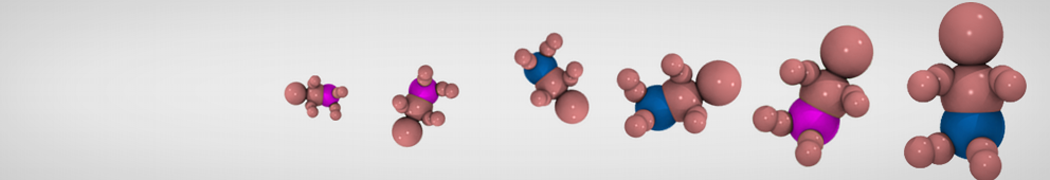
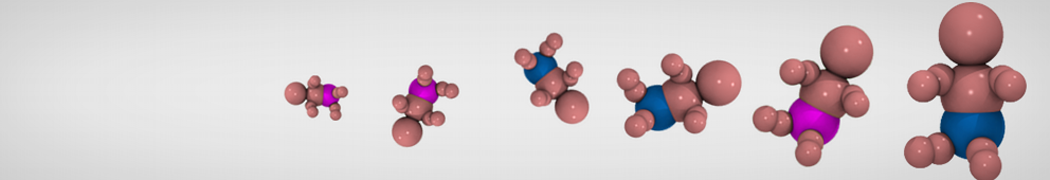
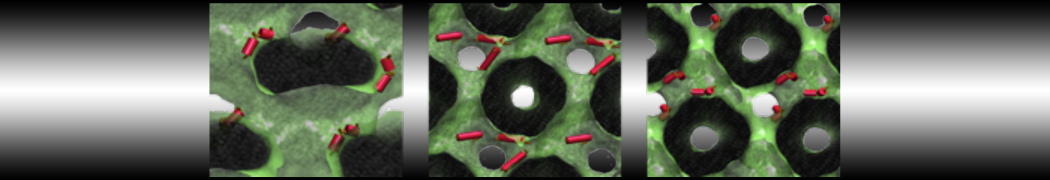
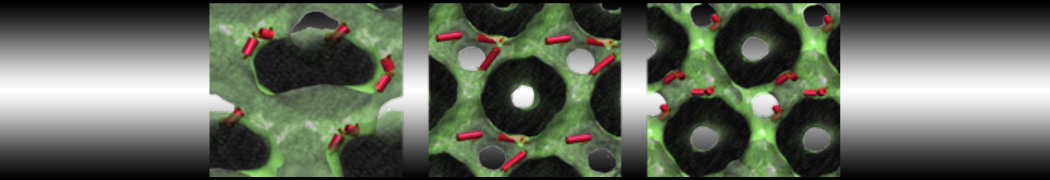
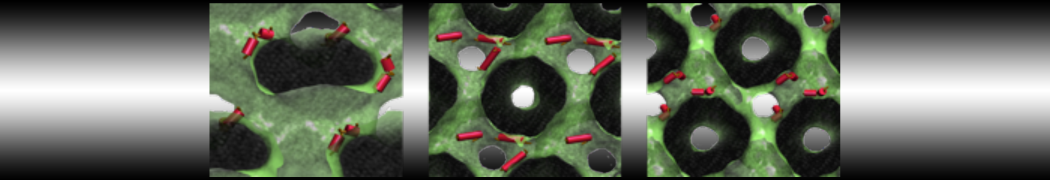
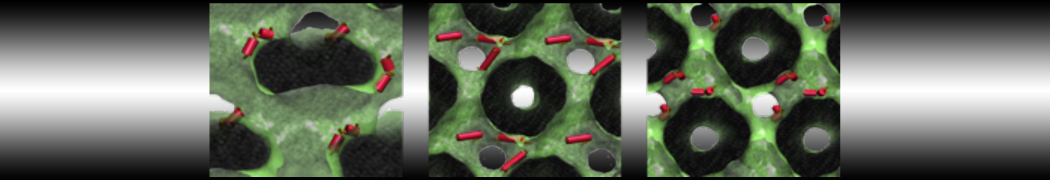
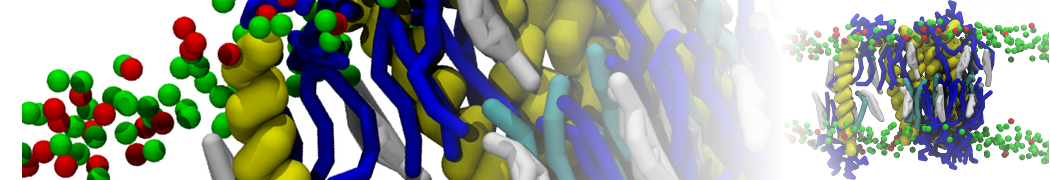
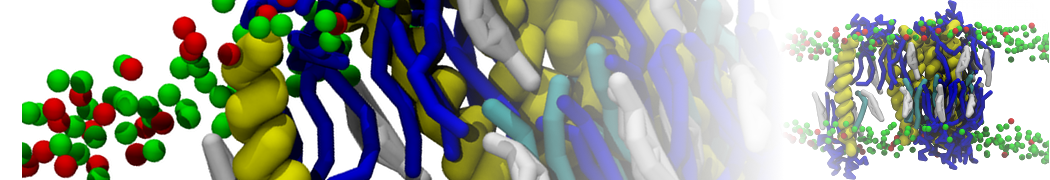
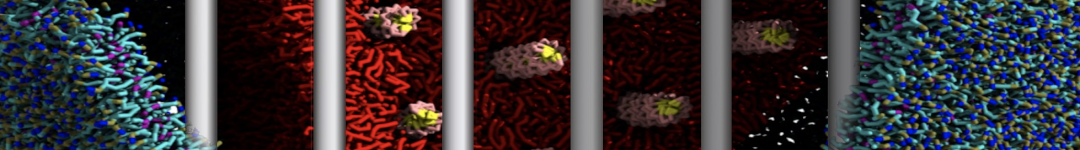
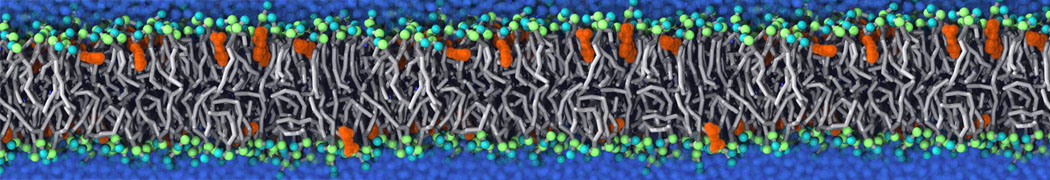
Hari S. Muddana, Homer H. Chiang, Peter J. Butler. Tuning Membrane Phase Separation Using Nonlipid Amphiphiles. Biophys J, 2012, 102:489-497
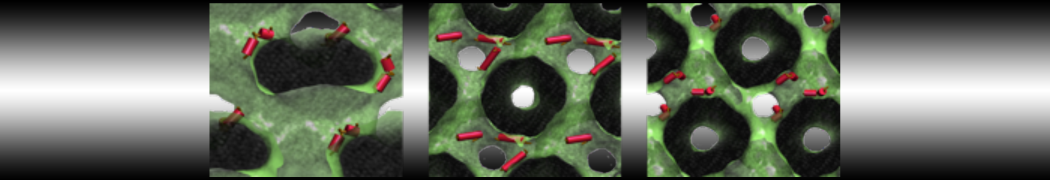
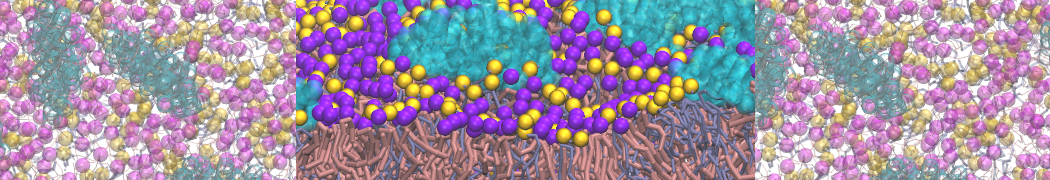
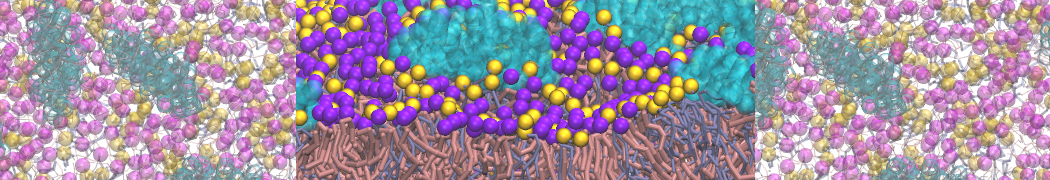
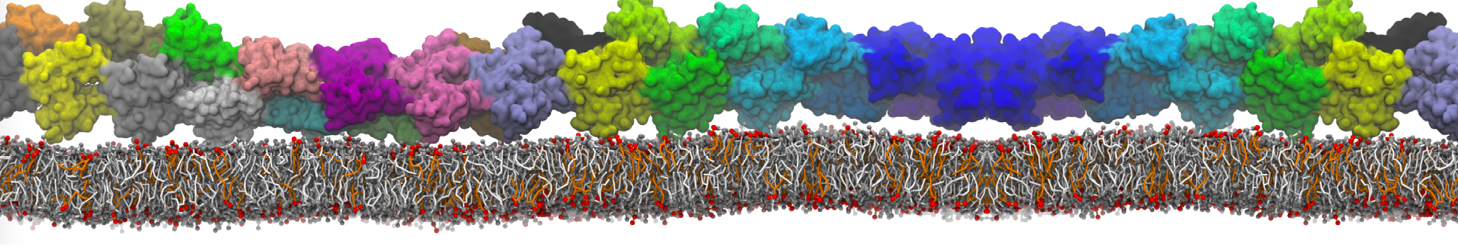
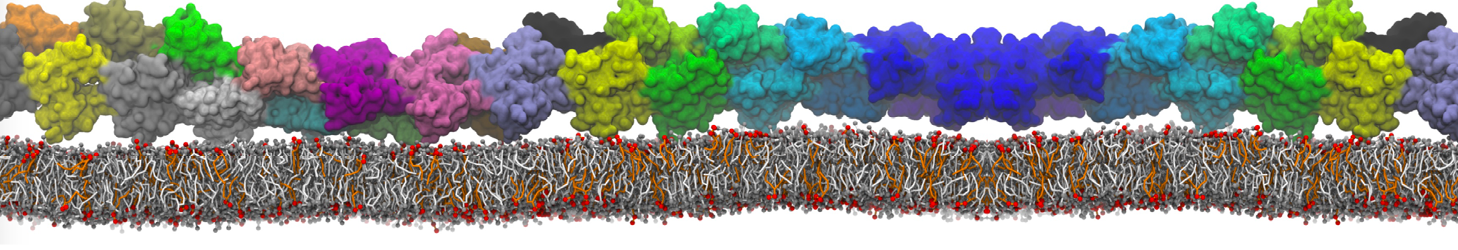
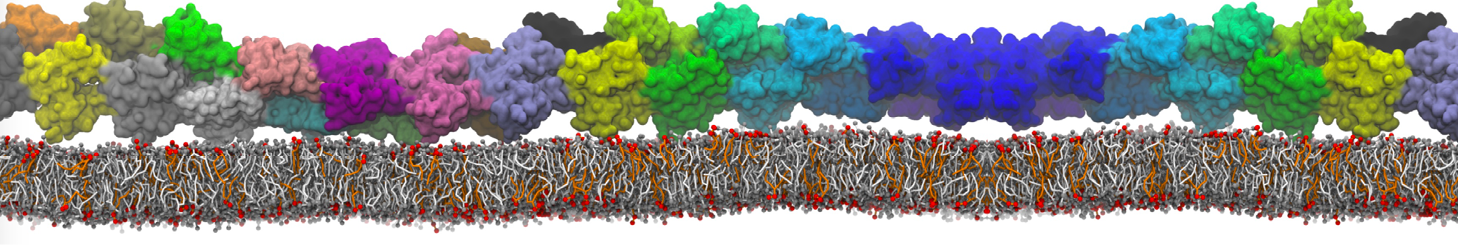
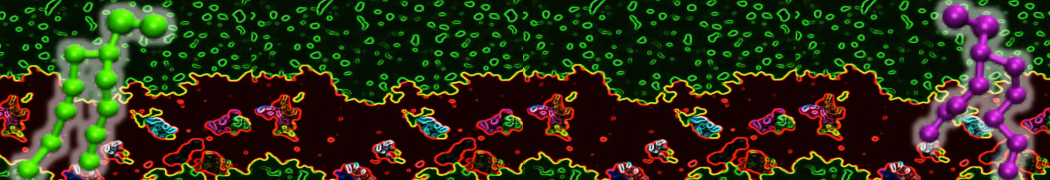
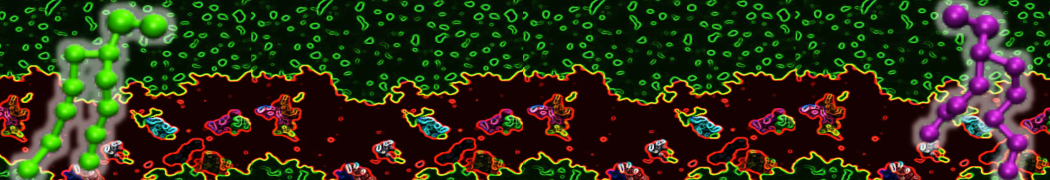
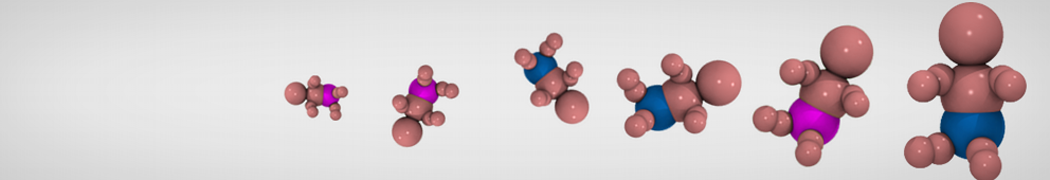
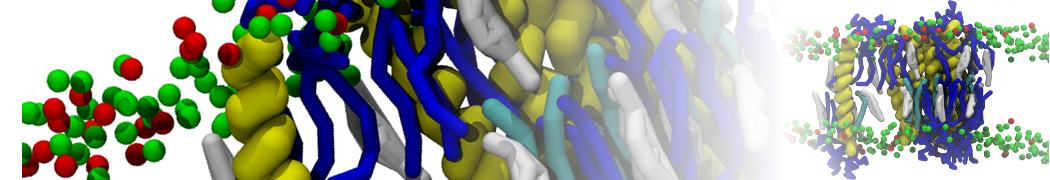
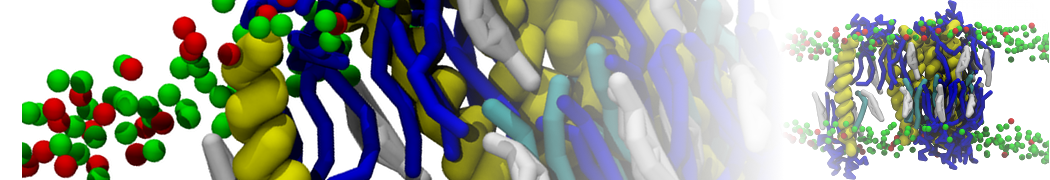
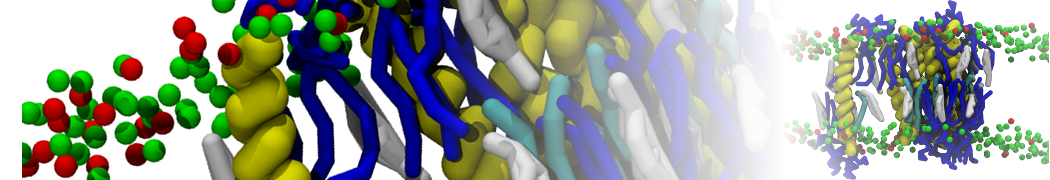
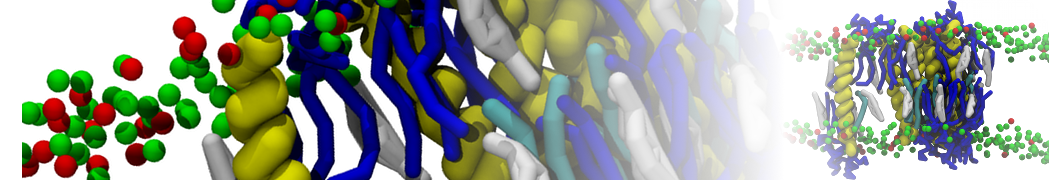
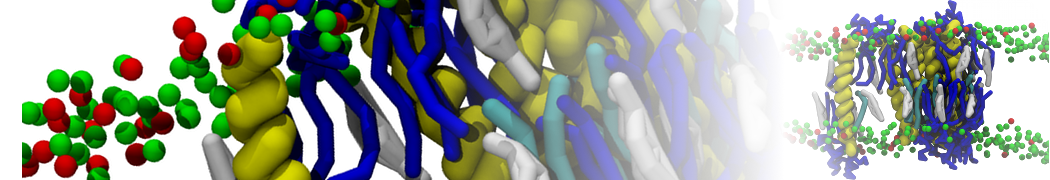
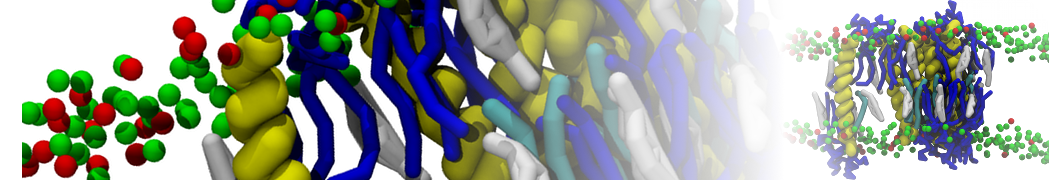
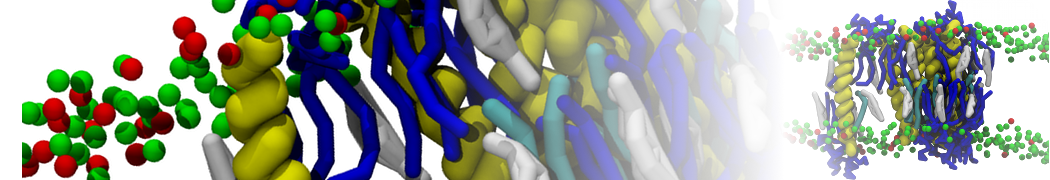
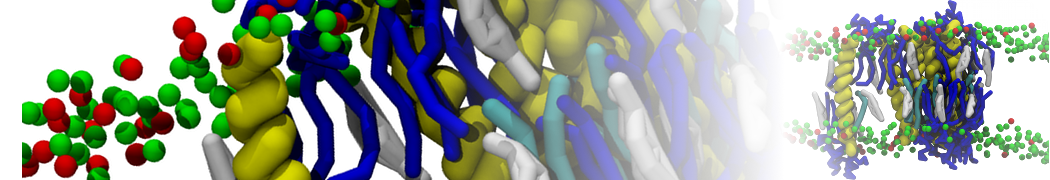
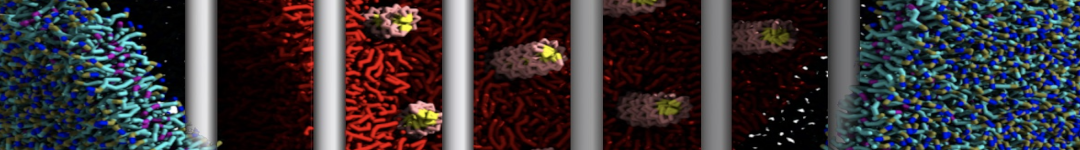
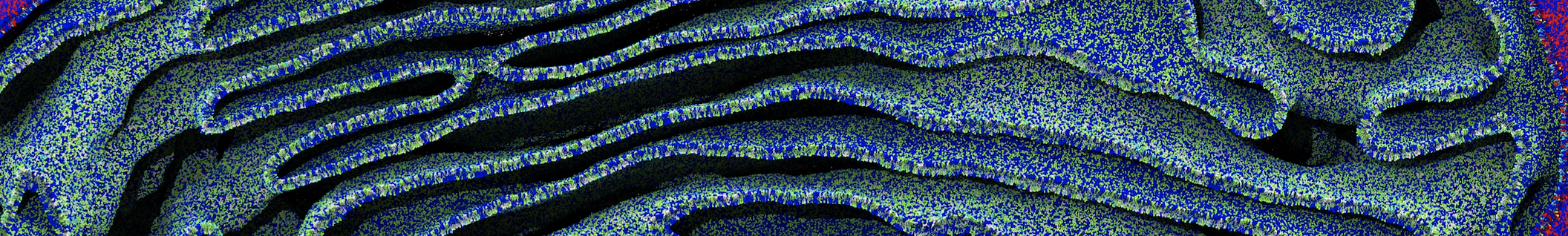
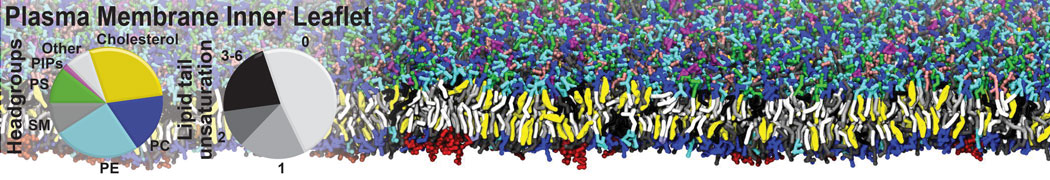
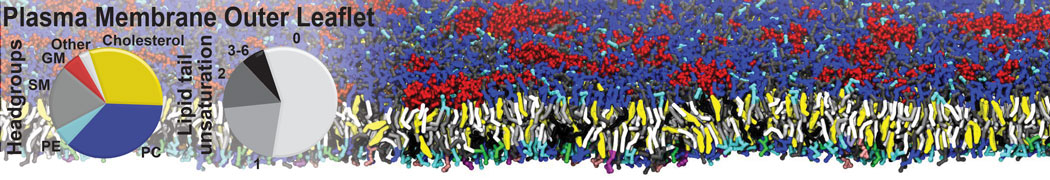
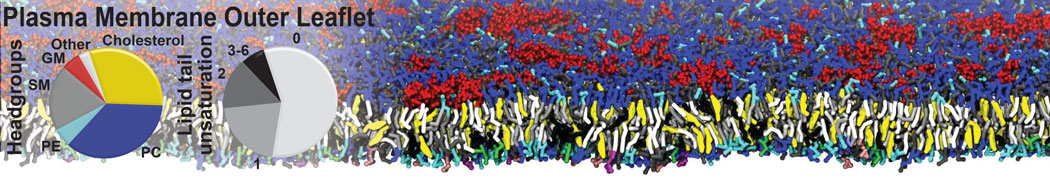
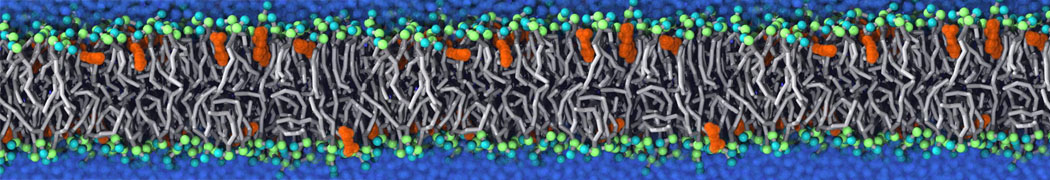
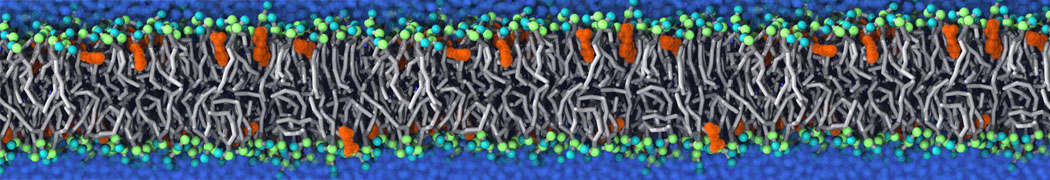
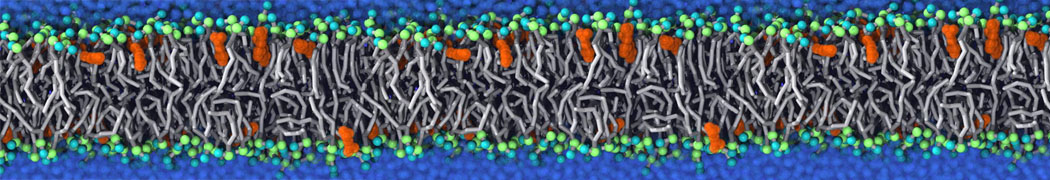
Lipid phase separation may be a mechanism by which lipids participate in sorting membrane proteins and facilitate membrane-mediated biochemical signaling in cells. To provide new tools for membrane lipid phase manipulation that avoid direct effects on protein activity and lipid composition, we studied phase separation in binary and ternary lipid mixtures under the influence of three nonlipid amphiphiles, vitamin E (VE), Triton-X (TX)-100, and benzyl alcohol (BA). Mechanisms of additive-induced phase separation were elucidated using coarse-grained molecular dynamics simulations of these additives in a liquid bilayer made from 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC) and 1,2-diundecanoyl-sn-glycero-phosphocholine (DUPC). From simulations, the additive's partitioning preference, changes in membrane thickness, and alterations in lipid order were quantified. Simulations showed that VE favored the DPPC phase but partitioned predominantly to the domain boundaries and lowered the tendency for domain formation, and therefore acted as a linactant. This simulated behavior was consistent with experimental observations in which VE promoted lipid mixing and dispersed domains in both gel/liquid and liquid-ordered/liquid-disordered systems. From simulation, BA partitioned predominantly to the DUPC phase, decreased lipid order there, and thinned the membrane. These actions explain why, experimentally, BA promoted phase separation in both binary and ternary lipid mixtures. In contrast, TX, a popular detergent used to isolate raft membranes in cells, exhibited equal preference for both phases, as demonstrated by simulations, but nonetheless, was a strong domain promoter in all lipid mixtures. Further analysis showed that TX increased membrane thickness of the DPPC phase to a greater extent than the DUPC phase and thus increased hydrophobic mismatch, which may explain experimental observation of phase separation in the presence of TX. In summary, these nonlipid amphiphiles provide new tools to tune domain formation in model vesicle systems and could provide the means to form or disperse membrane lipid domains in cells, in addition to the well-known methods involving cholesterol enrichment and sequestration.