Antimicrobial Peptides
- Details
- Last Updated: Friday, 14 July 2023 09:28
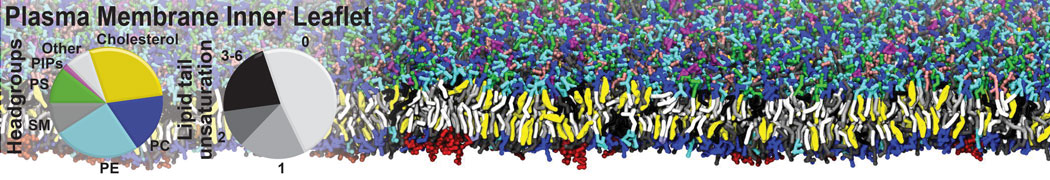
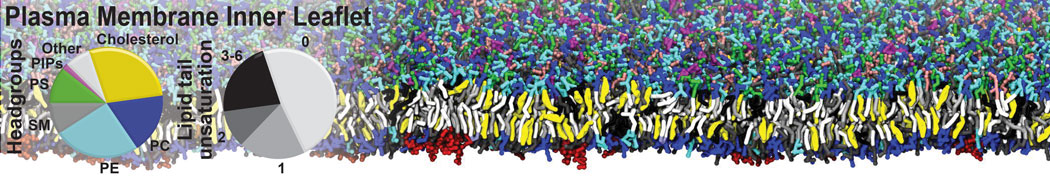
 General Purpose Coarse-Grained Force Field
General Purpose Coarse-Grained Force Field 











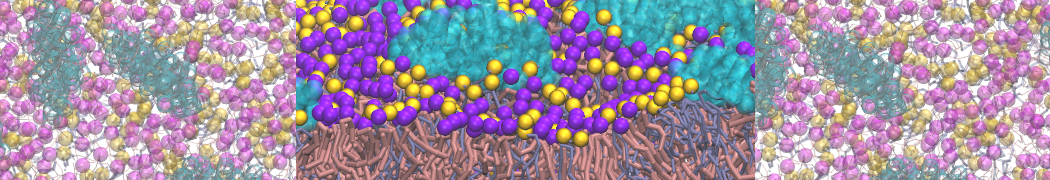
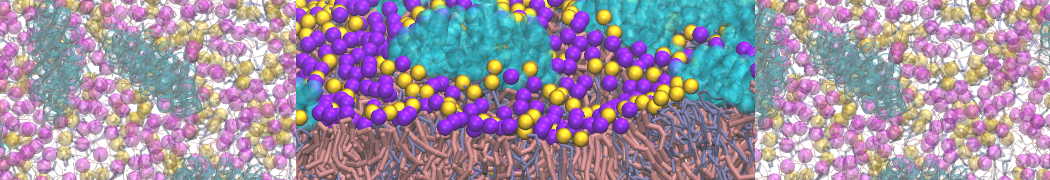
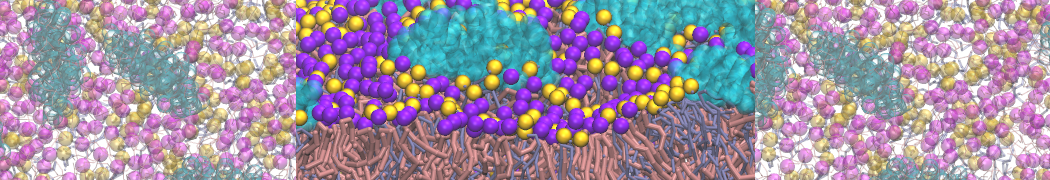
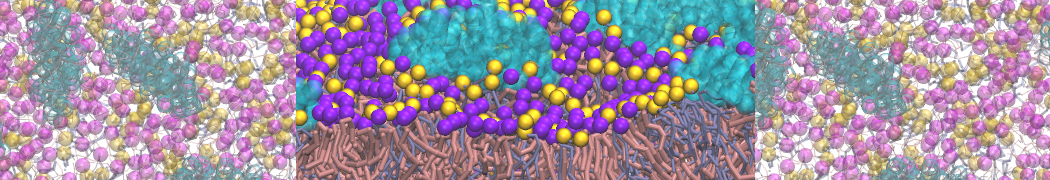
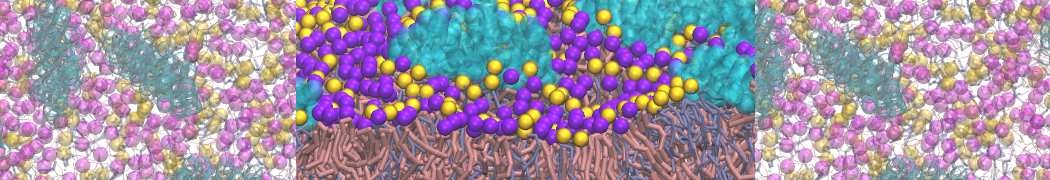




















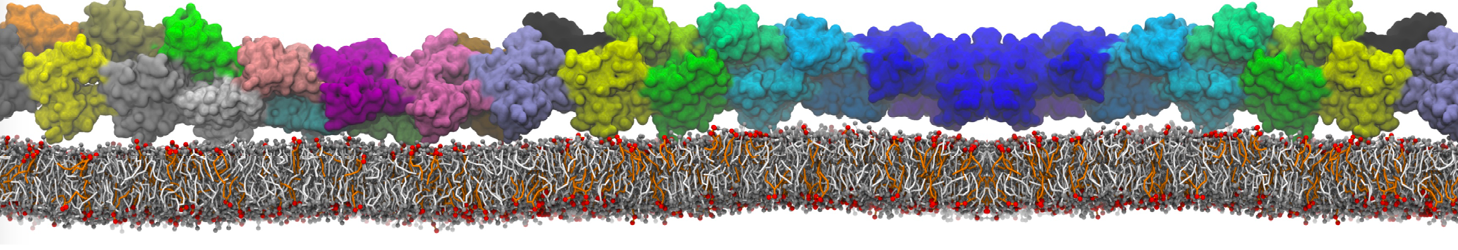
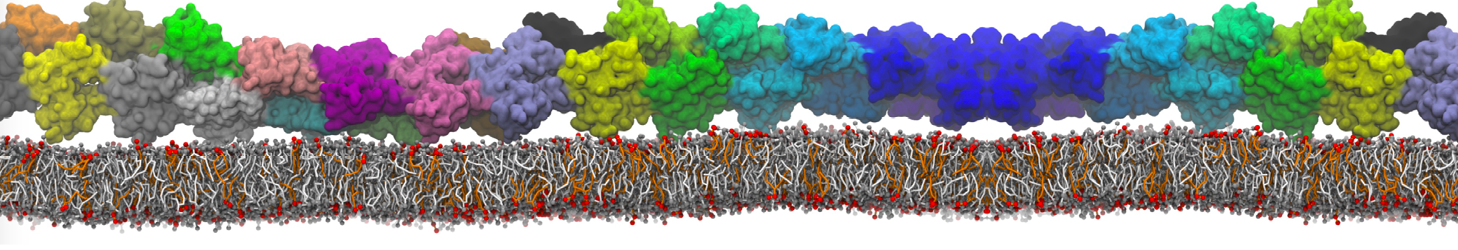










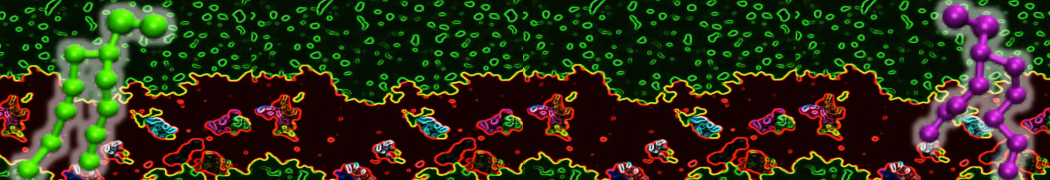
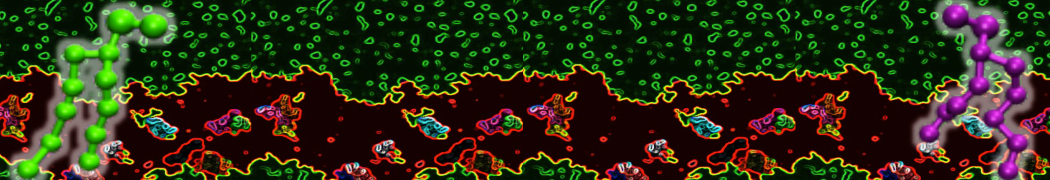
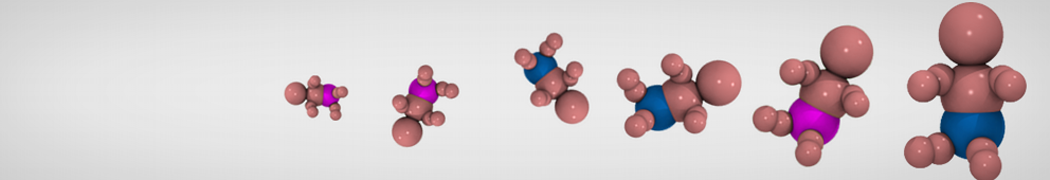
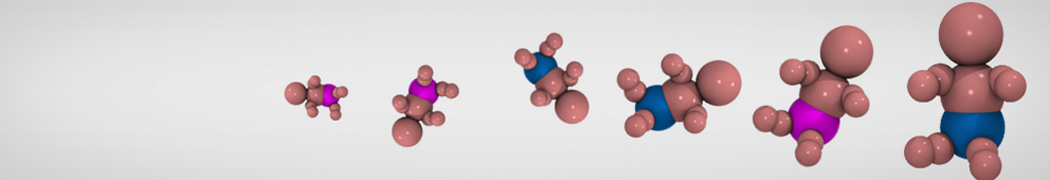
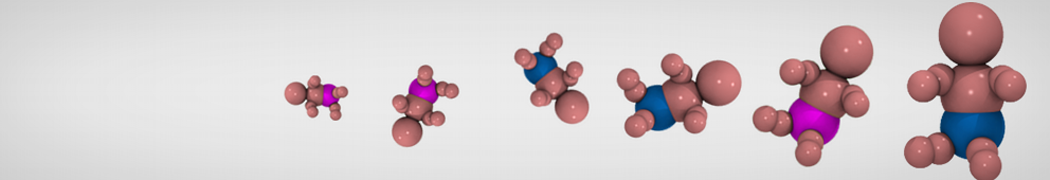
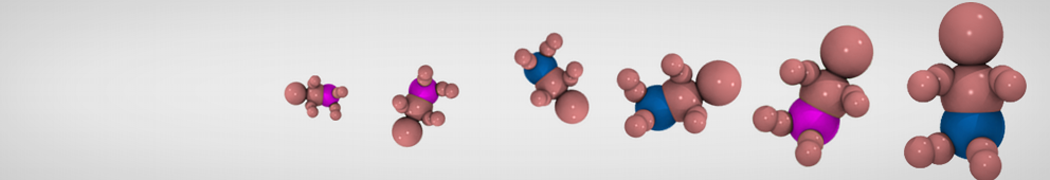
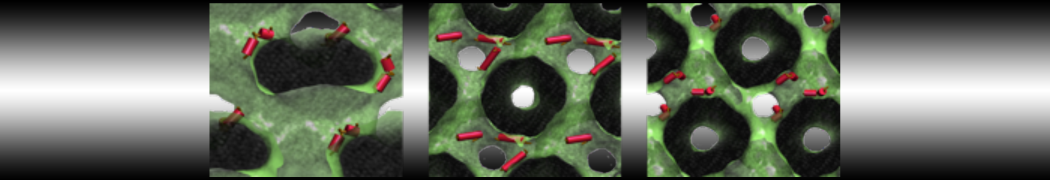
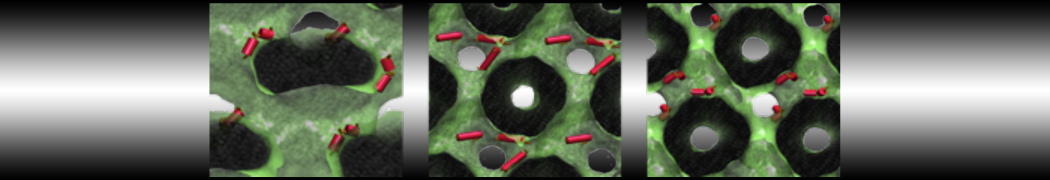
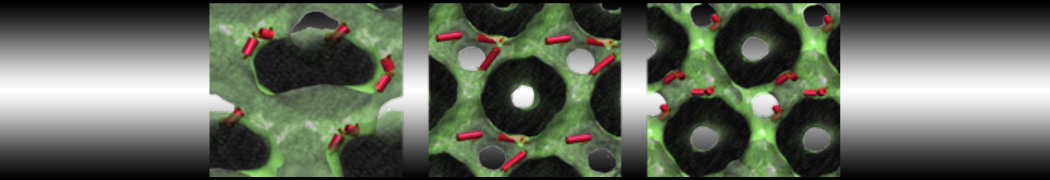
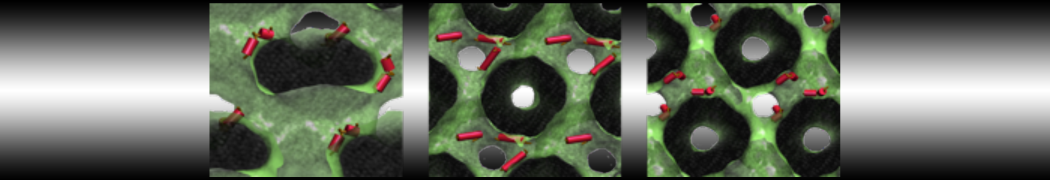
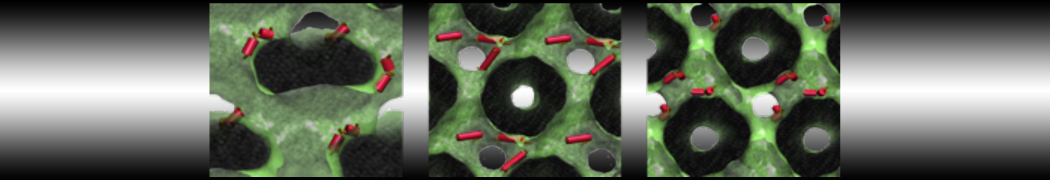
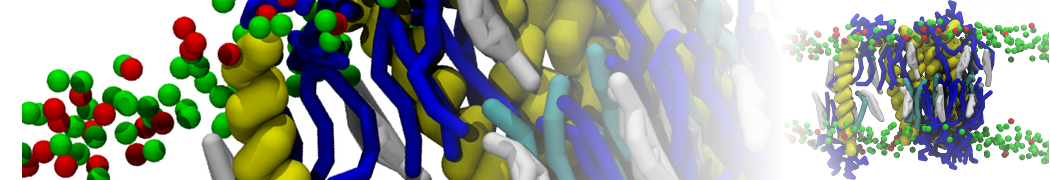
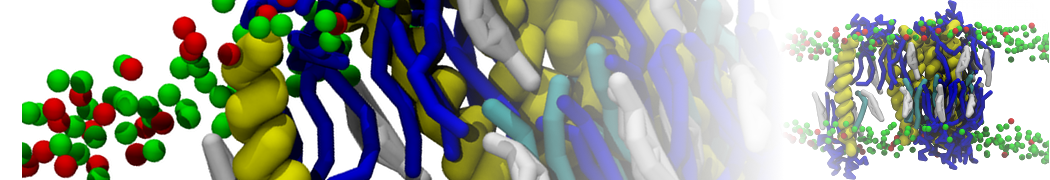
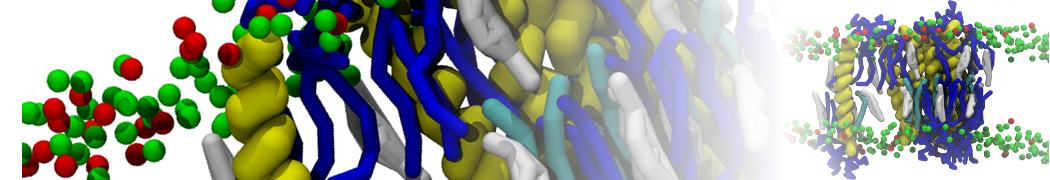
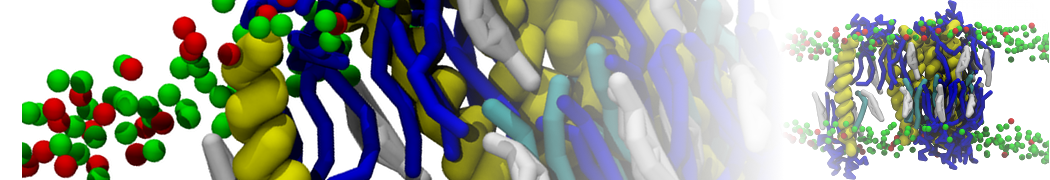
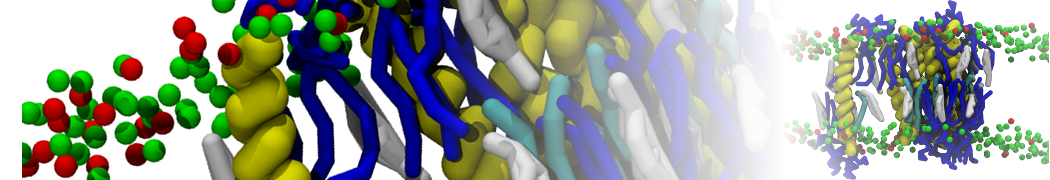
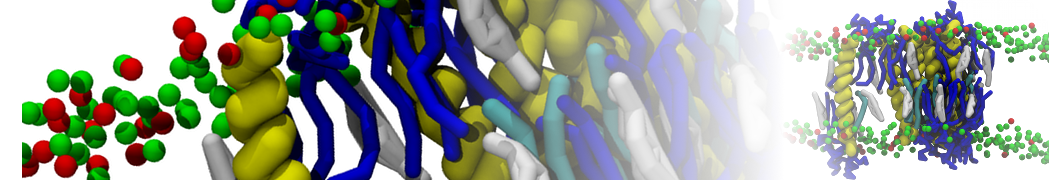
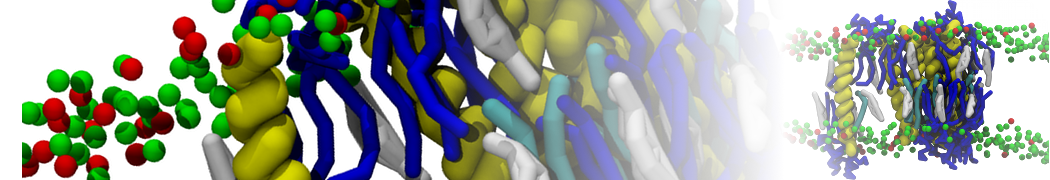
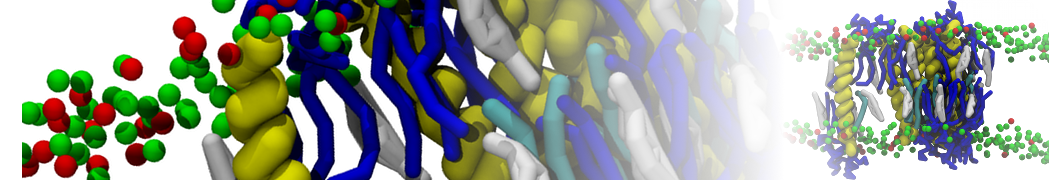
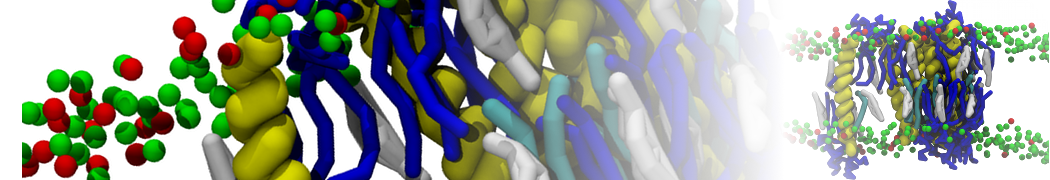
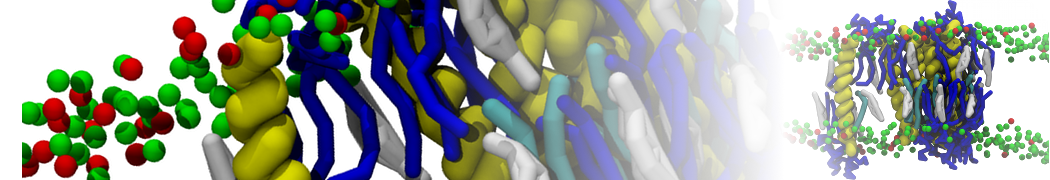











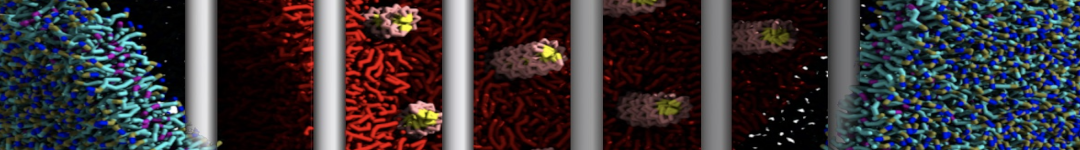
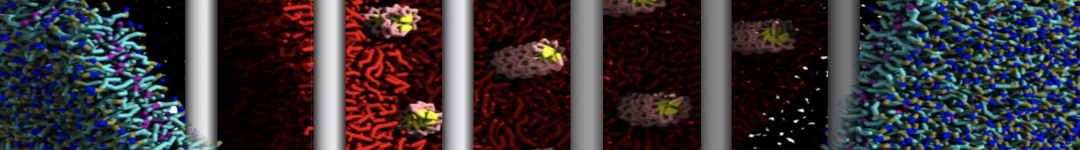
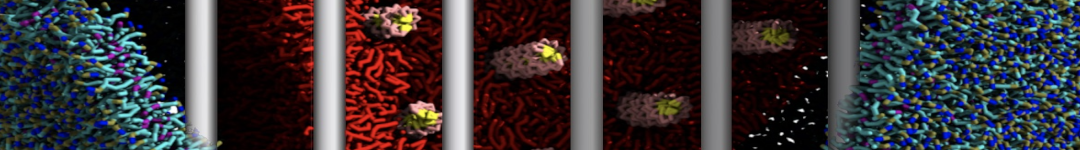
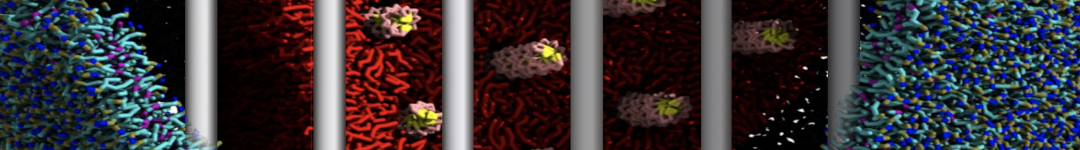
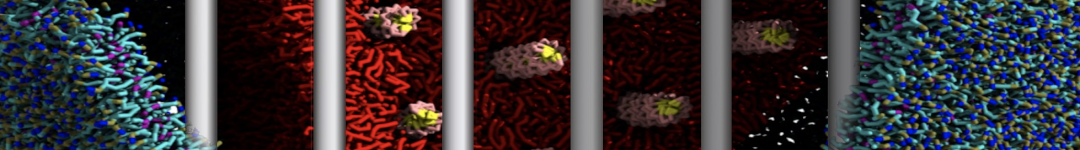






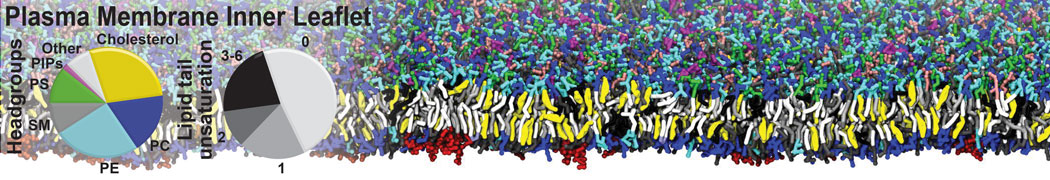
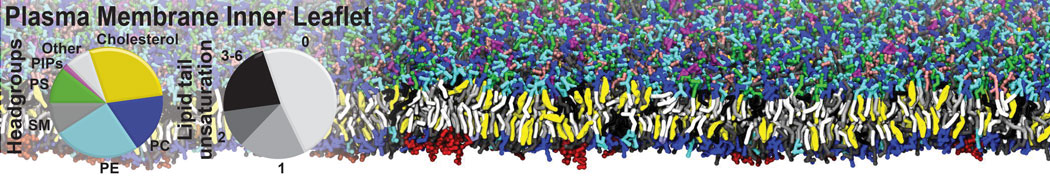
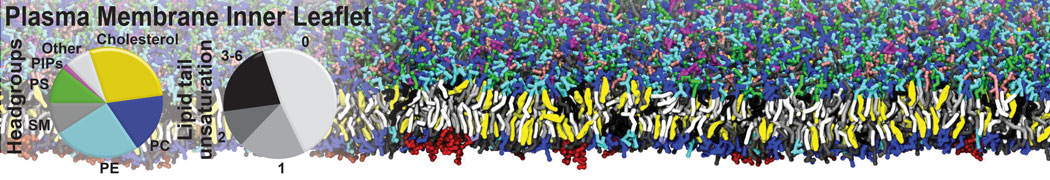
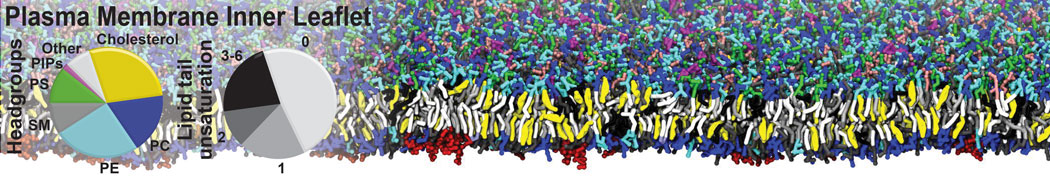















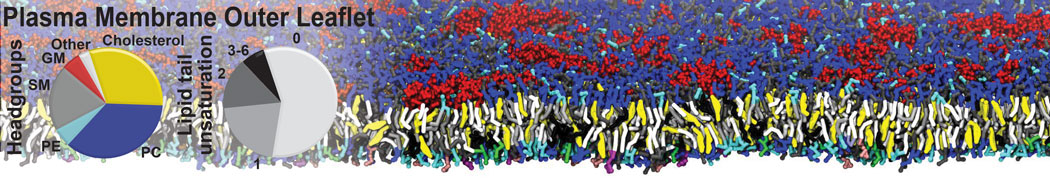
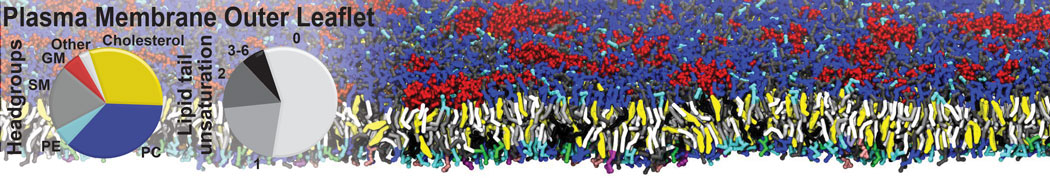
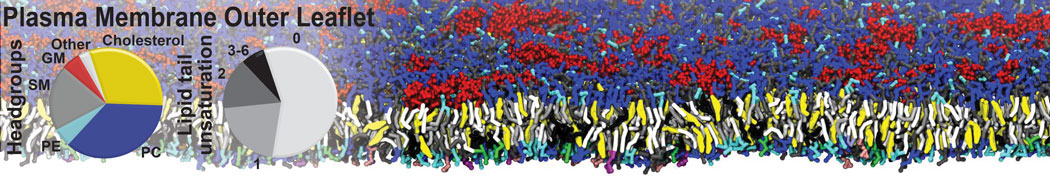
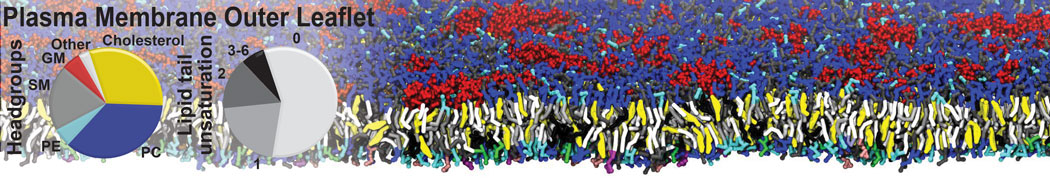
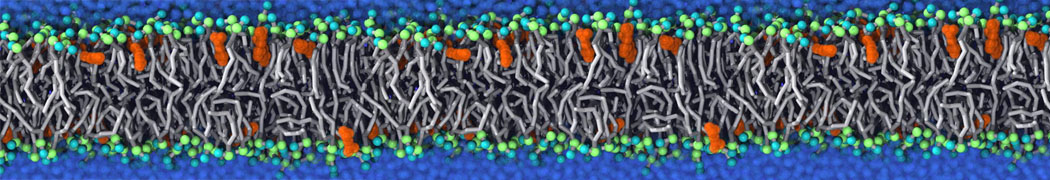
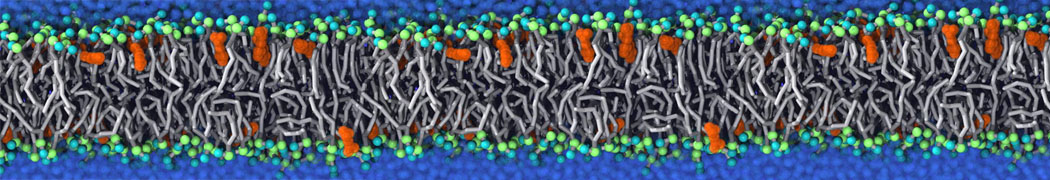
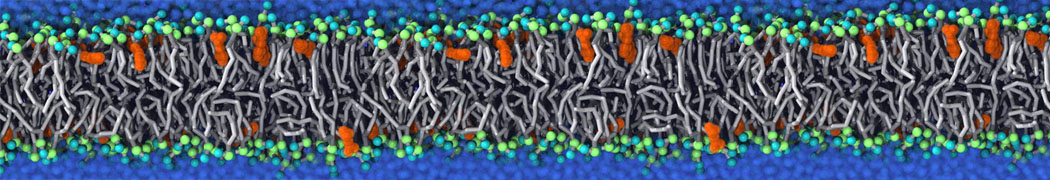
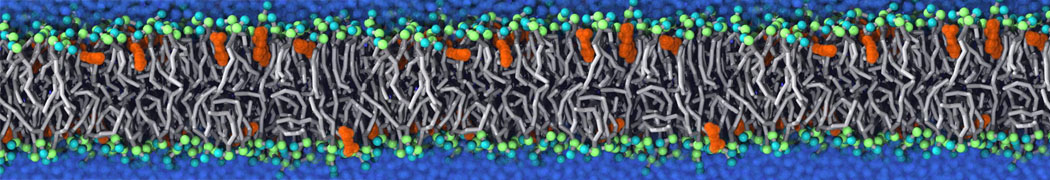
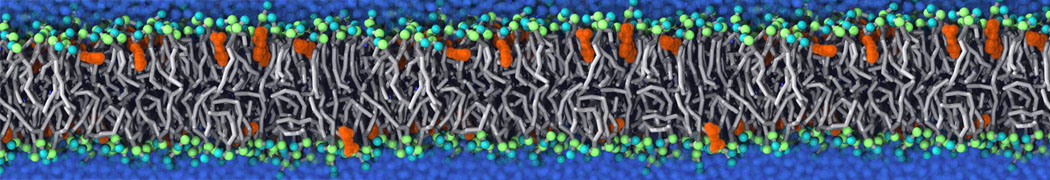
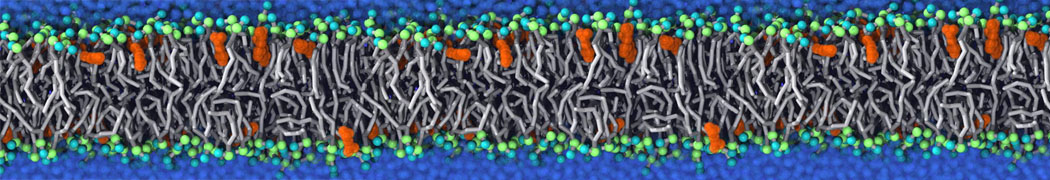
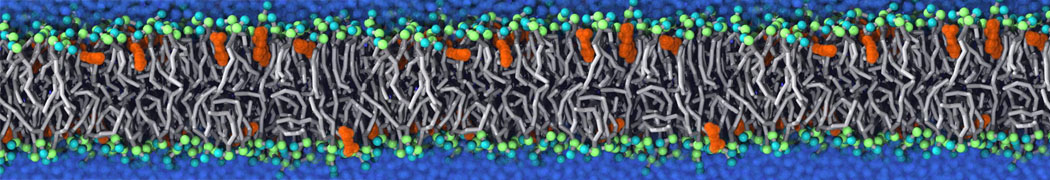
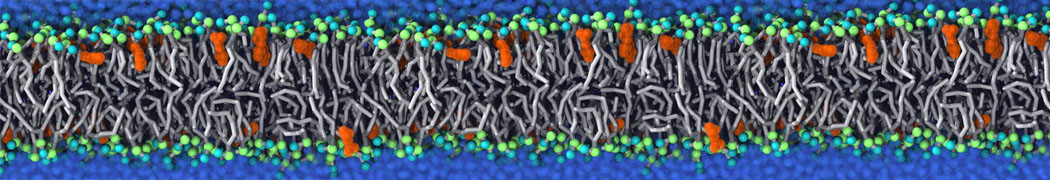
Synaptotagmin-1 may be a distance regulator acting upstream of SNARE nucleation
Geert van den Bogaart, Shashi Thutupalli, Jelger H Risselada, Karsten Meyenberg, Matthew Holt, Dietmar Riedel, Ulf Diederichsen, Stephan Herminghaus, Helmut Grubmüller & Reinhard Jahn. Nature Structural & Molecular Biology 18, 805–812 (2011)
Synaptotagmin-1 triggers Ca2+-sensitive, rapid neurotransmitter release by promoting interactions between SNARE proteins on synaptic vesicles and the plasma membrane. How synaptotagmin-1 promotes this interaction is unclear, and the massive increase in membrane fusion efficiency of Ca2+-bound synaptotagmin-1 has not been reproduced in vitro. However, previous experiments have been performed at relatively high salt concentrations, screening potentially important electrostatic interactions. Using functional reconstitution in liposomes, we show here that at low ionic strength SNARE-mediated membrane fusion becomes strictly dependent on both Ca2+ and synaptotagmin-1. Under these conditions, synaptotagmin-1 functions as a distance regulator that tethers the liposomes too far from the plasma membrane for SNARE nucleation in the absence of Ca2+, but while bringing the liposomes close enough for membrane fusion in the presence of Ca2+. These results may explain how the relatively weak electrostatic interactions between synaptotagmin-1 and membranes substantially accelerate fusion.
S.J. Marrink, J. Risselada, A.E. Mark. Simulation of gel phase formation and melting in lipid bilayers using a coarse grained model. Chem. Phys. Lip., 135:223-244, 2005.
The transformation between a gel and a fluid phase in dipalmitoyl-phosphatidylcholine (DPPC) bilayers has been simulated using a coarse grained (CG) model by cooling bilayer patches composed of up to 8000 lipids. The critical step in the transformation process is the nucleation of a gel cluster consisting of 20–80 lipids, spanning both monolayers. After the formation of the critical cluster, a fast growth regime is entered. Growth slows when multiple gel domains start interacting, forming a percolating network. Long-lived fluid domains remain trapped and can be metastable on a microsecond time scale. From the temperature dependence of the rate of cluster growth, the line tension of the fluid–gel interface was estimated to be 3 ± 2 pN. The reverse process is observed when heating the gel phase. No evidence is found for a hexatic phase as an intermediate stage of melting. The hysteresis observed in the freezing and melting transformation is found to depend both on the system size and on the time scale of the simulation. Extrapolating to macroscopic length and time scales, the transition temperature for heating and cooling converges to 295 ± 5K, in semi-quantitative agreement with the experimental value for DPPC (315 K). The phase transformation is associated with a drop in lateral mobility of the lipids by two orders of magnitude, and an increase in the rotational correlation time of the same order of magnitude. The lipid headgroups, however, remain fluid. These observations are in agreement with experimental findings, and show that the nature of the ordered phase obtained with the CG model is indeed a gel rather than a crystalline phase. Simulations performed at different levels of hydration furthermore show that the gel phase is stabilized at low hydration. A simulation of a small DPPC vesicle reveals that curvature has the opposite effect.
D. Sengupta, A. Rampioni, S.J. Marrink. Simulations of the C-subunit of ATP-synthase reveal helix rearrangements. Mol. Membr. Biol., 26:422-434, 2009.
The c-subunit of the enzyme, ATP synthase couples the proton movement through the a-subunit with its own rotation and subsequent rotation of the F1 ring to drive ATP synthesis. Here, we perform ms time-scale coarse-grained molecular dynamics simulations of the c-subunit to characterize its structure and dynamics. Two different helix-helix interfaces, albeit with similar interfacial characteristics, are sampled in the simulations. The helix-2 of the c-subunit monomer rotates around the axis of helix-1 bringing about a change in the interface. Previous models have also proposed such a change in the helix interface but postulated that helix-2 swivels around its own axis. Such large-scale changes in helix packing motifs have not been observed before. The helix-swirling persists even in the c-subunit ring but the dynamics is much slower. The cooperative behavior in the ring appears to stabilize a conformation less-populated in the monomer. Analyzing the stability of the c-subunit ring, it was found that six lipid molecules are necessary to fill the central cavity of the ring. These lipid molecules were not aligned with the surrounding bilayer but protruded towards the periplasmic side. The characterization of the monomer and ring presented in this work sheds light into the structural dynamics of the c-subunit and its functional relevance.
L.V. Schäfer, S.J. Marrink. Partitioning of lipids at domain boundaries in model membranes. Biophys. J., in press, 2010.
Line-active molecules (“linactants”) that bind to the boundary interface between different fluid lipid domains in membranes have a strong potential as regulators of the lateral heterogeneity that is crucial for biological membranes. Here, we use molecular dynamics simulations in combination with a coarse-grain model that retains near-atomic resolution to identify lipid species that can act as linactants in a model membrane that is segregated into two lipid domains of different fluidity. We find that certain hybrid saturated/unsaturated chain lipids can bind to the interface and lower the line tension, whereas cone-shaped lysolipids have a less pronounced effect. This study demonstrates that line-active behavior can be predicted from molecular dynamics simulations.
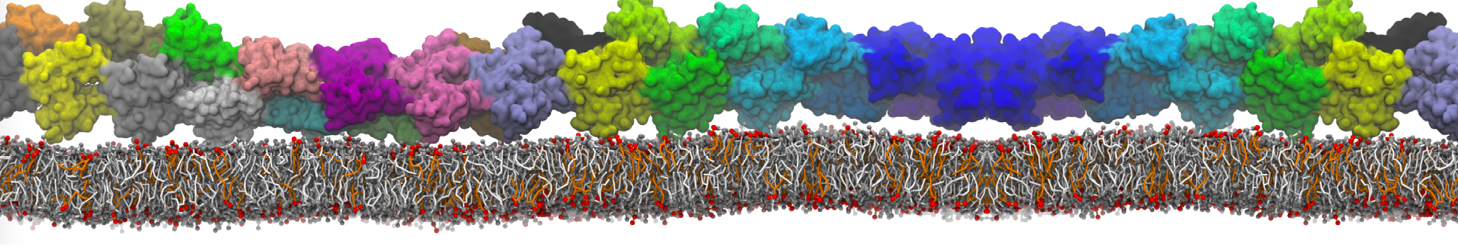
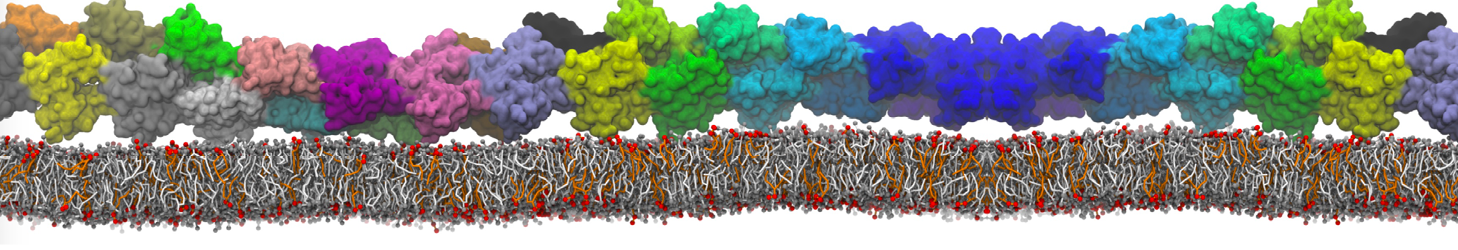
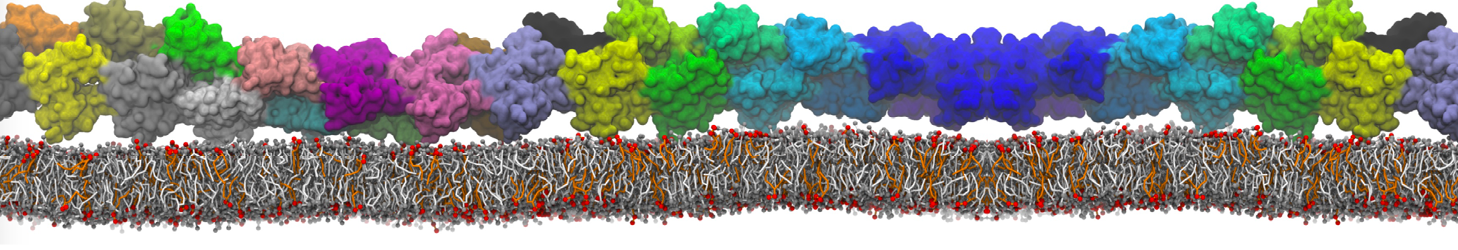
"Mechanism of Substrate Shuttling by the Acyl-Carrier Protein within the Fatty Acid Mega-Synthase" by
Fatty acid mega-synthases (FAS) are large complexes that integrate into a common protein scaffold all the enzymes required for the elongation of aliphatic chains. In fungi, FAS features two independent dome-shaped structures, each 3-fold symmetric, that serve as reaction chambers. Inside each chamber, three acyl-carrier proteins (ACP) are found double-tethered to the FAS scaffold by unstructured linkers; these are believed to shuttle the substrate among catalytic sites by a mechanism that is yet unknown. We present a computer-simulation study of the mechanism of ACP substrate-shuttling within the FAS reaction chamber, and a systematic assessment of the influence of several structural and energetic factors thereon. Contrary to earlier proposals, the ACP dynamics appear not to be hindered by the length or elasticity of the native linkers, nor to be confined in well-defined trajectories. Instead, each ACP domain may reach all catalytic sites within the reaction chamber, in a manner that is essentially stochastic. Nevertheless, the mechanism of ACP shuttling is clearly modulated by volume-exclusion effects due to molecular crowding and by electrostatic steering toward the chamber walls. Indeed, the probability of ACP encounters with equivalent catalytic sites was found to be asymmetric. We show how this intriguing asymmetry is an entropic phenomenon that arises from the steric hindrance posed by the ACP linkers when extended across the chamber. Altogether, these features provide a physically realistic rationale for the emergence of substrate-shuttling compartmentalization and for the apparent functional advantage of the spatial distribution of the catalytic centers.